Древние рыбообразные (Agnatha и ранние Gnathostomata) представляют собой критически важный этап в эволюции позвоночных, охватывающий период от силура до девона.
Класс Agnatha (бесчелюстные), из которого до наших дней дожили только миноги и миксины, в древности был представлен многочисленными вымершими группами, такими как бесщитковые (Anaspida), костнощитковые (Osteostraci) и разнощитковые (Heterostraci).
Эти существа, такие как Cephalaspis и Pteraspis, характеризовались внешним костным или дентиновым панцирем и отсутствием парных плавников, что указывает на придонный образ жизни и фильтрационное питание.
Рис. 1. Акулообразная рыба Climatius с 7 боковыми шипами, соответствующими, очевидно, парным плавникам.
Содержание страницы
К этому классу принадлежат по крайней мере четыре подкласса. О. Абель (1924) считает наиболее примитивным подклассом силурийских бесщитковых (Anaspi da).
У этих своеобразных животных тело было одето несколькими рядами косо расположенных покровных пластинок.
Жаберная коробка поддерживалась 8—10 рядами жаберных дуг; на голове за единственным носовым отверстием располагалось от верстие для теменногоглазка.
Хвост был снабжен неравномерным плавником; имелся анальный плавник. Костнощитковые подкласса Osteostraci характеризовались большим костным головным щитком.
Лучше других представителей известен Cephalaspis, который встречался от верхнего силура до верхнего девона.
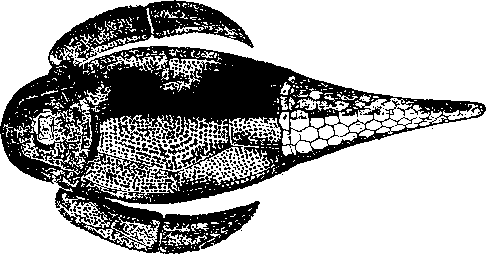
Весьма примитивный подкласс составляют силуро-девонские разнощитковые (Heterostraci).
Они отличались широким сжатым спереди туловищем; хвост был относительно суженный, заканчивающийся гетероцеркальным или чаще резко вырезанным плавником.
Глаза были расставлены широко и помещались по бокам головы, рот был расположен в ее нижнем отделе. Жаберные отверстия не обнаружены.
Кожа была покрыта жесткими чешуями из дентина; у некоторых видов на теле были большие пластины, например, на переднем отделе туловища.
У девонского Drepanaspis голова по внешнему виду не отличается от туловища, и весь передний отдел туловища необыкновенно расширен: хвост узкий, покрытый чешуей.
На дорзальной и вентральной сторонах туловища находится по большой пластине. Рот беззубый.
У девонского Pteraspis сплошной спинной щит был вытянут на голове в узкий носовой выступ; был деференцирован значительный брюшной щит.
Тело имело вид торпеды.
Каждый небольшой глаз был охвачен особым кольцом.
Эволюция круглоротых в настоящее время остается еще весьма неясной. Этот класс, как полагают Грегори и Осборн, очень рано, вероятно, в силуре, отделился от первоначального ствола рыбообразных существ (см. выше).
И вследствие приспособлений к паразитическому существованию дегенерировал, выработав в то же время ряд специальных признаков своей организации.
К предкам круглоротых многие относят Palaeospondilus gunni из девонских отложений Шотландии.
Рис. 2 . Ptenchtys millen.
Эволюционное развитие настоящих рыб (Pisces). В основу класса настоящих рыб, по новейшим данным, ставят подкласс крылатых (Antiarchi), так как у них найдены челюсти.
К Antiarchi принадлежат рыбы, имеющие наружный панцырь, состоящий из крупных пластин.
Передние конечности имеют вид очень крупных ластов и слагаются как бы из отдельных частей. Из среднего девона известен своеобразный Pterichtys milleri (рис. 2).
Antiarchi представляют специализированный подкласс древних настоящих рыб Pisces).
Представители наиболее примитивных акул известны из девона; диференцировка ствола поперечноротых, вероятно, началась еще с силура (Грегори и Осборн).
Из девонских акул особенно интересна Cladoselache fyleri из девона Северной Америки. Примитивно устроенные плавники этой акулы описаны нами выше.
Со сравнительно-анатомической стороны крайне интересна нижнедевонская Climatius (рис. 1) с рядами мелких брюшных плавников, что показывает возможность филогенетического развития парных плавников из цельной боковой складки.
Из современных акул наиболее древним можно считать сем. Cestracionidае, известное еще с каменноугольного периода.
В рецентной фауне сохранился только один вид Hetero-dontus philippi, распространенный в Тихом океане у австралийских берегов.
Скаты — специализированная ветвь акул.
Без сомнения, они обособились от общего ствола акул и приспособились к определенным условиям существования.
Эти рыбы появляются лишь с верхнеюрского периода.
Большой интерес представляет история эволюции двоякодышащих рыб.
Наибольшего расцвета этот подкласс достигает в девонской и каменноугольной эпохах; затем начинается обеднение видового состава двоякодышащих.
В то же время если проследить историю описываемых рыб, то можно увидеть постепенный процесс специализации наряду с регрессом некоторых морфологических признаков.
В самом деле, древнейшие представители двоякодышащих, как нижнедевонский Dipterus valenciennesii (рис. 3).
Отличались гетероцеркальным хвостом, двумя спинными плавниками, небольшим анальным и двумя группами ластовидных парных плавников.
Жаберная крышка этой рыбы была широка и несла слой ганоина; тело покрывали ганоидные чешуи.
У следующей формы, Dipterus macropterus, хвост острее, задний спинной плавник больше; у верхнедевонской Scaumenacia замечается еще большее увеличение и продвижение вперед спинного плавника.
У последующих, более новых, видов хвостовой плавник непосредственно переходил в спинной, и вере-теновидное тело рыб приобретает дифицеркальный хвост.
Анальный плавник сливается с хвостовым. У Neoceratodus чешуя резко утоньшается, жаберная крышка сокращается в размерах.
У Protopterus и Lepidosiren наблюдается заметная редукция жабер и жаберных крышек, также численно сокращаются и уменьшаются в размерах чешуйки кожи; конечности приобретают характер жгутов.
Рис. 3. Эволюция признаков от Dipterus до Lepidosiren.
Последовательно снизу вверх: Dipterus valenciennesii, Dipterus macropterus, Scaumenacia curta, Phaneropleron andersoni, Uronemus lobatus, Neoceratodus forsteri, Protopterus annectens, Lepidosiren para-doxa.
При обзоре последовательной эволюции двоякодышащих мы констатируем прогрессивное развитие некоторых признаков, например, образование легких, изменение структуры кровеносной системы.
В этом можно видеть процесс филогенетического ускорения и приобретения особенностей амфибиальных животных.
С другой стороны, в развитии дифицеркального хвоста из более специализированного гетероцеркального сказывается определенный регресс.
Несомненные предки хрящевых ганоидов известны из нижнего лейаса. К ним принадлежит беззубый и голый с не вполне окостеневшим скелетом Chondrosteus acipenseroides.
Кистеперые имеют своих предков еще в девоне. К ним принадлежат Osteolepis macrolepidotus (из среднего девона Шотландии) и знаменитый p. Rhizodus (из карбона), интересный структурой плавников, могущих служить как бы прототипом пятипалой конечности.
Рис. 4. Схема эволюционного развития рыб.
Настоящие костистые рыбы (Teleostei) представляют высшую ступень специализации и развития вторичных признаков, сменивших первичные и обусловивших большую стойкость организма в жизненной конкуренции.
Остатки костистых рыб, имевших костный скелет и кожный покров из тонких чешуи, встречаются в мезозое, начиная с мела; более ранние предки найдены, в среднем триасе (Leptolepidae) (рис. 4).
Для ясности соотношений различных систематических категорий даем классификацию вымерших и современных рыб, указывая главнейшие группы:
А. Рыбообразные (ICHTYOMORPHI)
Класс I. Круглоротые (Cyclostomata).
Класс II. Панцырные (Placodermi)
Подкласс 1. Бесщитковые (Anaspida)
2. Разнощитковые (Heterostraci)
3. Костнощитковые (Osteostraci)
Б. Рыбы собственно (PISCES)
{kind=link}
{kind=link}